景天酸代谢途径植物的光合研究
背景资料:
景天酸代谢途径(crassulacean acid metabolism pathway,CAM途径),指生长在热带及亚热带干旱及半干旱地区的一些肉质植物(早发现在景天科植物)所具有的一种光合固定二氧化碳的附加途径。具有这种途径的植物被称为CAM植物。在其所处的自然条件下,气孔白天关闭,夜晚张开。它们具有此途径,既维持水分平衡,又能同化二氧化碳。途径的特点是:在夜间细胞中磷酸烯醇式丙酮酸(PEP)作为二氧化碳接受体,在PEP羧化酶催化下,形成草酰乙酸,再还原成苹果酸,并贮于液泡中;白天苹果酸则由液泡转入叶绿体中进行脱羧释放二氧化碳,再通过卡尔文循环转变成糖。所以这类植物的绿色部分的有机酸特别是苹果酸有昼夜的变化,夜间积累,白天减少。淀粉则是夜间减少(由于转变为二氧化碳接受体PEP)白天积累(由于进行光合作用的结果)。已发现许多科植物如龙舌兰科、仙人掌科、大戟科、百合科、葫芦科、萝藦科,以及凤梨科具有此途径。一般说CAM植物是多汁的,但也有不是多汁的。多汁植物也并不都是CAM植物。这类植物是通过改变其代谢类型以适应环境,由于该途径的特点造成光合速率很低(3~10毫克CO2·分米-2·小时-1),故生长缓慢,但能在其它植物难以生存的生态条件下生存和生长。如果说C4植物是空间上错开二氧化碳的固定和卡尔文循环的话,那景天酸循环就是时间上错开这两者。
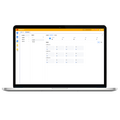
图1仙人掌科和凤梨科的植物
具有景天酸代谢途径的植物光合作用的研究:
CAM植物因为具有这特殊光合途径,这也就使得其光合作用的测定和其他植物的测定有很大不同。
1 CAM植物叶片往往形状各异,肉质,叶片厚度较大,如上图的仙人掌等。(目前上的光合仪叶室主要是为作物如玉米、小麦、水稻、大豆等设计,因此也就很难用于CAM植物的叶片)
2光合仪(便携性光合作用测定仪)的原理是根据光合作用的总反应式:
CO2+ 2H2O*+ 4.69kJ → (CH2O) +O*2+ H2O
通过红外线CO2气体分析仪测定CO2浓度的变化来计算光合速率。而CAM植物白天气孔关闭,不和外界进行气体交换,因此,用光合仪是不能测定CAM植物的光合速率。这也是很多用户用光合仪测定有些植物光合一直在0附近的根本原因。
3 CAM植物光合速率很低,测定必须要用比其他作物更精密的仪器来测定。
基于以上特点要想测定CAM植物的光合速率我们必须另辟新径。前几章讲过,对于用光合仪测定不了的叶片不规则的植物材料可以用氧电极测定。
方法一:用氧电极测定CAM植物的光合速率,
根据光合作用的总反应式:CO2+ 2H2O*+ 4.69kJ → (CH2O) +O*2+ H2O
氧电极测定光合速率和光合仪测定的光合速率不同(详见“光合仪和氧电极测定光合速率的区别及优缺点”),氧电极是通过氧气的释放测定光合速率的,由于反应在液相体系中进行,就消除了气孔对气体释放的影响。而且氧电极精确度非常高,不易受到CAM植物光合速率低的影响。
可以用来测定CAM植物光合速率的氧电极有:Chlorolab-2和Chlorolab-3型(Hansatech公司,英国)
方法二:用CO2分析仪在晚上多点测定群体CO2的吸收,来估计净光合积累
CAM植物是通过时间把二氧化碳的固定和卡尔文循环分开,晚上打开气孔吸收CO2,因此我们就可以在晚上用CO2分析仪估测。由于CAM植物的光合速率一般较低,因此即使晚上吸收的CO2比较少,因此我们可以测定群体对CO2的吸收,这样就可以用光合仪配有的群体同化室进行测定,群体同化室采用密闭式气路就可以(CAM光合速率较低),晚上多点测定,像白天测定日变化一样,分几个点测定CO2的吸收,这样可以估算CAM植物的光合积累。
光合仪一般都配有群体同化室,如CIRAS-2配CPY-2、CPY-3型同化室(PP Systems公司,美国)
综上所述,我们在使用光合仪测定光合速率的时候一定要注意,如果自己研究的实验材料是龙舌兰科、仙人掌科、大戟科、百合科、葫芦科、萝藦科,凤梨科中的一种,就一定要注意该材料是否是CAM植物,如果是CAM植物一般就要选择氧电极进行测定光合速率。氧电极是可以测定光合仪不能测定的材料,如藻类,植物的特殊部位等。













![聚酰胺粉 [柱层析用,高分离性能] 60-100目/80-120目/100-200目](https://p-06.caigou.com.cn/135x120/2024/7/2024071513085253637.jpg)